

The game I play is a very interesting one.
It’s imagination in a tight straitjacket.
Like Richard Feynman’s game of science, evolution is stuck in a straitjacket. It is driven by chance. But evolution is not free to explore every path.
Take, as an example, the evolution of organism size. While it seems like there are many routes to bigness, I propose that there is fundamentally only one: sociality. In the march towards ever-larger organisms, there have been three major revolutions. All of them involved the merger of previously autonomous organisms into a new communal creature. I call this route to bigness ‘size through sociality’. It is a tale 4 billion years in the making.
The drive towards sociality, I argue, is a response to a basic feature of geometry. As objects get larger, their volume grows faster than their surface area. This fact of space causes problems for harvesting energy. It requires that big organisms harness and distribute energy on a limited surface-area budget. The easiest way to solve this problem, it seems, is to merge existing structures. Hence the evolution of bigger life is deeply connected to the evolution of sociality.
The evolution of ‘big’ is also connected to human culture.
Modern human institutions may represent a new transition in the evolution of life — a transition from massive organisms to supermassive superorganisms. But as with the rest of life, this evolution occurs in a straitjacket. The size distribution of human institutions seems to follow the same pattern as the size distribution of other organisms. In fact, it is an extension of this pattern, upping the size of life to new proportions.
In this light, human cultural evolution may be a variation on an old theme: size through sociality.
The tyranny of geometry
As I child, I loved playing with toy cars. I made the little vehicles jump over great distances, usually with the gleeful hope that they might explode. But the toys always took the beating with ease. When I imagined doing the same stunt with life-sized cars, however, I knew that they couldn’t withstand the punishment. But I didn’t know why.
Today I do. It’s because the size of an object changes how it behaves. The reason owes to a simple feature of geometry. As objects get larger, their volume grows faster than their area. This fact affects the objects’ properties.
Let’s use a sphere to illustrate. As you make a sphere bigger, its volume grows with the cube of its radius:
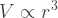
This volume determines the mass of the sphere — the amount of stuff that must be held together. The strength of the material, however, is proportional to cross-section area. And that grows with the square of the radius:
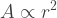
So we have a mismatch between volume (mass) and cross-section area (strength). Because volume grows faster than area, objects get weaker as they get larger. That’s why toy cars can take more beating than real cars. Relative to their mass, toy cars are proportionally stronger.
The puddle-of-guts problem
This strength-scaling problem applies not only to cars, but to life. It’s why big animals are build differently than small animals. To support their bulk, elephants need proportionally thicker limbs than do mice.
The strength-scaling problem is also why a Godzilla-sized creature will never exist. At 100 meters tall, the beast would be so massive that its bones and muscles couldn’t support it. As Neil deGrasse Tyson quips, “Godzilla would collapse under his own weight into a puddle of guts.”
The starvation problem
Long before any animal reached the puddle-of-guts limit, it would run into a more pressing problem. It would starve.
A simple way to understand the problem is to treat an organism as a ‘reverse sun’. The sun radiates energy outward through its surface. An organism does the reverse. It absorbs energy though its surface and ‘radiates’ it inward.
In both cases, the flow of energy is limited by surface area. This fact poses problems for big organisms. Their ability to harvest energy is proportional to their surface area, which (assuming a spherical organism) grows with the square of their radius:

But as we saw above, the organism’s volume grows with the cube of its radius.
So again we have a mismatch — another tyranny of geometry. As an organism gets bigger, its mass grows faster than its ability to feed itself. So long before a Godzilla-sized creature pushed the limits of material strength, it would probably starve to death.
The evolution of ‘big’
Now that we’ve covered the basic limits to organism size, let’s review the evolution of bigness. Life started small and, over 4 billion years, gradually got larger. The results now surround us — a diversity of life that varies from minuscule bacteria to monumental blue whales.
It’s tempting to think that the tyranny of geometry manifests itself only at the top of life’s scale, keeping blue whales from growing larger still. But that’s false. At several landmark stages, evolution had to overcome barriers to size.
Figure 1 gives a birds-eye view of this problem-solving process. I’ve plotted here something called the ‘biomass spectrum’ — the relation between organism size and abundance. As organisms get larger, they become more rare.1
As organisms get larger, they also change form. The various taxa of life occupy different niches of size. In other words, blue whales are not enlarged bacteria. Nor are bacteria shrunken blue whales.

The long march towards bigness (visible as we move from left to right in Figure 1) didn’t involve simply scaling up existing features. It required evolving new structures that sidestepped the tyranny of geometry. That’s why life has different niches of size. Bacteria are not small because they choose to be. Bacteria are small because they have to be.
Although the forces of evolution produced a wild diversity of life, the march towards greater size was underwritten by only 3 major transitions. Life started with the microscopic prokaryotic cell, which was the sole type of organism for 2 billion years. Then, in the first major transition of evolution, prokaryotes went social, forming the eukaryotic cell. Life got larger. In the next major transition, eukaryotes went social, forming multicellular organisms. Life got larger still. In the final transition (not shown in Figure 1), multicellular organisms went social, forming superorganisms. Life became monumentally large. Box 1 summarizes the story.
Box 1: Major transitions in the size of life
Transition 1: prokaryotic cell ⟶ eukaryotic cell
Transition 2: eukaryotic cell ⟶ multicellular organisms
Transition 3: multicellular organisms ⟶ superorganisms
What unites these transitions is that they achieved greater size by increasing sociality. Evolution took previously autonomous organisms and stuck them together, making them ‘social’.
Why the common solution? Probably because evolution is lazy. Rather than build something from scratch, evolution prefers to tinker with what exists.
With ‘size through sociality’ in mind, let’s look at the evolution of ‘big’.
In the beginning was the cell
How did life start? The short answer is that no one knows. But we do have some interesting ideas.
In his book The Vital Question, biochemist Nick Lane argues that life’s Promethean invention was the cell. His reasoning boils down to thermodynamics. Life is what physicists call a ‘dissipative structure’. It is a structure that arises to dissipate energy. Dissipative structures, of course, are not unique to life. They occur everywhere that energy flows. Interestingly, they often take the form of cells.
When you boil water, for instance, you’ll see little cells emerge. Under the right conditions, they can look like the picture on the left of Figure 2. These structures are called convection cells, and they arise spontaneously when thermal energy passes through a liquid.

Although convection cells are not ‘alive’, they bear an eerie resemblance to the cells of living creatures — for instance to the bacteria on the right of Figure 2. Nick Lane thinks this resemblance is not a coincidence. Both types of cells (dead and living) do the same thing: they dissipate energy. But they do it in different ways.
Convection cells dissipate a thermal gradient (a heat difference within the liquid). Living cells, in contrast, dissipate a chemical gradient. In this regard, life works much like a battery. Living cells maintain a chemical potential across a membrane. From this potential comes a flow of energy, which makes life possible.
What do cells and their membranes have to do with organism size? It boils down to the tyranny of geometry.
Across all life on Earth, metabolism happens along membranes — a surface area. But that membrane has to supply energy for all the stuff inside it — a volume. Since surface area grows more slowly than volume, cells run out of energy as they get larger. That is why life started small. And that is why much of it remains small.
The prokaryotic revolution: life in the microscopic
When the first cells emerged 4 billion years ago, they were likely simple — little more than membranes filled with fluid. To see such simple cells, we need not look into the deep past. These cells surround us. They’re in us. They’re called prokaryotes, and they are the most numerous form of life.
Prokaryotes are a type of single-celled organism distinguished by their lack of internal structure. Today, we know them as bacteria (and the lesser-known archaea).2 These simple creatures are the oldest form of life, and the smallest. While other organisms have become massive, prokaryotes have remained microscopic. Why?
Nick Lane thinks the answer is simple: it’s the tyranny of geometry.
Prokaryotes derive energy by synthesizing ATP along their cell membrane. So their ability to harness energy scales with surface area. But their need for proteins scales with cell volume. (Selective pressure, Lane argues, keeps prokaryotes from slimming down their genome.) The result is that as prokaryotes grow larger, they run out of energy for protein synthesis. Or in simpler terms, they starve.
So while prokaryotes have been spectacularly successful, they are doomed to be small.
The eukaryotic revolution: prokaryotes go social
While prokaryotes are stuck in their microscopic niche, other life has escaped to become gigantic. How?
The answer, Nick Lane argues, is by evolving mitochondria.
All large organisms are composed of eukaryotic cells — cells that have mitochondria. In these cells, ATP isn’t produced along the outer membrane (as it is with prokaryotes). Instead, eukaryotes rely on their mitochondria to synthesize ATP. It’s a clever solution to the tyranny of geometry.
Like prokaryotes, mitochondria still make ATP along their membrane. But as the host eukaryotic cell gets larger, the mitochondria don’t grow. They become more numerous, thus maintaining a high surface-area-to-volume ratio. Importantly, mitochondria also have a lean genome, which means that little energy must be devoted to protein synthesis.
So if mitochondria are the solution to the prokaryotic size problem, where did they come from? The answer appears to be ‘size through sociality’.
The eukaryotic cell is not one cell, but many. It is an archaean cell that plays host to symbiotic bacteria living inside it. Over eons, the bacteria shed most of its DNA and evolved into today’s mitochondria. This symbiosis, Nick Lane observes, gave eukaryotic cells ‘multibacteria power’, but with a fraction of the protein costs. As a result, eukaryotic cells have copious energy at their disposal:
Large eukaryotes such as amoebae have more than 200,000 times the energy per gene than a giant scaled-up bacterium.(Nick Lane, 2015)
By becoming hosts for symbiotic bacteria, eukaryotic cells freed themselves of the energy constraints faced by autonomous prokaryotes. Or put another way, a group of rogue prokaryotes solved their size problem by going social.
The multicellular revolution: eukaryotes go social
When prokaryotes went social (creating eukaryotes) it allowed a significant increase in the size of life. Unlike bacteria, which are microscopic, some single-celled eukaryotes are visible to the naked eye. Still, we never find amoebas the size of blue whales. But why not?
The answer (again) is the tyranny of geometry.
Recall the puddle-of-guts problem — the fact that strength grows more slowly than mass. This problem limits the size of single-celled creatures.
Cells are essentially sacks of liquid held together by a membrane. When the cell is small, a thin membrane will do the job of holding the cell together. That’s good, because the membrane is also the cell’s transport system — a highway for taking in nutrients and expelling waste. This highway only works when the membrane is thin, and hence, semi-permeable. A big cell would need a proportionally thicker membrane. But that would make it impermeable, gumming up the nutrient highway.
Large cells also have a difficult time circulating materials within their insides. That’s because cells (for the most part) circulate materials passively, through diffusion. That works well when you are small. But as volume grows, diffusion becomes painfully slow. (As an example, it takes tens of thousands of years for a photon of light to diffuse from the center of the sun to the surface.) The fact that diffusion does not scale well is why all large animals have an active circulatory system.
Faced with limits to cell size, life hit upon the same solution as before: size through sociality. But this time, instead of one cell absorbing another, a group of cells clumped together and began living communally. Multicellular life was born.
In fact, multicellularity was born not once, but many times. So powerful was the multicellular solution that it was arrived at independently by many forms of life. After the multicellular revolution, groups of communal cells grew trillions of times larger than their single-celled counterparts. As they did, multicellular life came to dominate Earth’s biomass.
The invertebrate world
A general rule in evolution seems to be this: whatever comes first remains the smallest and most numerous organism. Prokaryotes exemplify the rule. They were the first form of life. And today, they remain the smallest and most numerous organism.
Among multicellular organisms, the equivalent of prokaryotes are invertebrates. They were the first form of multicellular life, and they remain the smallest and most numerous animal. So what’s holding down invertebrate size?
On land it’s quite clear that the problem is the tyranny of geometry. Land invertebrates either go without a skeleton, or wear it on their outsides. Either way, they are subject to the puddle-of-guts problem, whereby strength scales more slowly then volume. At a certain size, you simply must have a skeleton to hold yourself together. And wearing it on your outside is inefficient and heavy. So vertebrates win the size war.
Invertebrates are also subject to the starvation problem, whereby surface area limits the ability to harvest energy. The problem for invertebrates is that their breathing system is largely passive. Insects, for instance, don’t circulate oxygen through their blood. Instead, they rely on a passive tracheal system that brings air directly to tissue. Because the system relies on diffusion, it does not scale well. As insects get more massive, they become starved of oxygen.3
In the sea, in contrast, limits to invertebrate size are likely not physical so much as they are evolutionary. The problem is the pesky vertebrates. It’s in regions where vertebrates are sparse that invertebrates grow enormous. In the fish-free deep, for instance, the 1000 pound colossal squid thrives. But where fish are abundant, vertebrates tend to be smaller.
So while sea-fairing invertebrates could be much larger, they seem unable to compete with fish. The reason is probably very simple: without a spine, it’s difficult to swim fast. So the advantage goes to fish, who eat spineless invertebrates for breakfast.4
The reign of mammals
Today, warm-blooded birds and mammals are the dominant large animals. But this was never destined to be true. A hundred million years ago, mammals were mouse-size creatures living in the shadow of the dinosaurs. When the dinosaurs were serendipitously wiped out, mammals filled the size void.
Why didn’t reptiles fill the void?
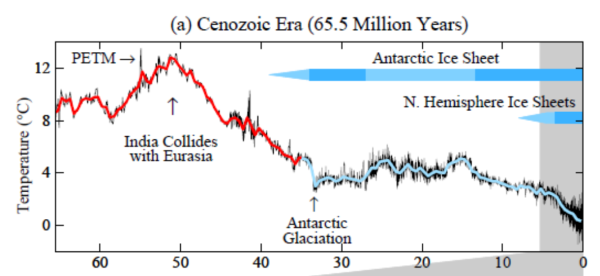
The problem may have been climate change. About 50 million years ago, the Earth started to cool (Figure 3). That may have posed problems for cold-blooded reptiles, who could not regulate their body temperature. Cold-blooded reptiles can get quite large (i.e. crocodiles), but they do so in the tropics where it is warm. Mammals and birds, however, can thrive in much colder temperatures.5
Lest we (mammals) be too triumphant about our position at the top of life’s pyramid, we should realize that thermoregulation comes with risks. For one thing, it locks us into being fairly large.
As size decreases, mammals face their own tyranny of geometry. Maintaining body temperature requires balancing heat production (a function of volume) with heat loss (a function of surface area). As mammals get smaller, their volume decreases faster than their surface area, causing excess heat loss. Below about 2 grams, land mammals cannot maintain their body temperature. So we’re stuck above that size.6
Another problem with thermoregulating is that it makes you vulnerable to extreme heat. When the wet-bulb temperature gets above 37°C, humans die. Given that we’re rapidly warming the Earth, this heat-death vulnerability doesn’t bode well for our future.
The limits of multicellularity
Back to the march towards bigness. The multicellular revolution allowed life to grow to mammoth proportions. It’s worth putting the scale in perspective. The world’s largest animal, the blue whale, is about 100 million-trillion times more massive than the smallest bacteria. Incredible.
Could multicellular life grow larger still?
The answer is probably yes, but only a by little. The problem, again, is the tyranny of geometry. Larger animals have more mass to feed and less surface area with which to feed it. At a certain point, the animal becomes so large that it can’t find enough food to feed itself.
Consider the blue whale. It makes its living by roaming the seas in search of enormous troves of krill. When it finds such a trove, it gorges by the ton. The blue whale’s extreme size makes it a highly efficient krill-eating machine. But the strategy works only when the krill is concentrated. If you took the same quantity of krill and spread it thinly across the ocean, the blue whale couldn’t survive.7
My guess is that the blue whale is pushing the limits of multicellularity. The Earth doesn’t have a dense enough food source to support an animal many times bigger. So if life wants to far surpass blue-whale size, it will need a new strategy — a new way of being social.
The superorganism revolution: multicellular organisms go social
The problem with multicellularity is that it requires that communal cells touch one another. This leads to the starvation problem. When an organism gets extremely large, the cells have too much mass and too little surface area for eating.
If life were to sidestep this limit, it would need a way of being social that did not require touching. Imagine a Godzilla-sized animal whose components could spread out over the Earth to harvest food. Were such an organism to exist, it would have the best of both worlds. It would have the benefits of size, but lack the constraints caused by the tyranny of geometry.
How would you build such a creature? The answer is that you build it not out of cells, but out of multicellular organisms. You create animals that are so social that they function like a single organism — a superorganism.
For much of the 20th century, the idea of superorganisms remained controversial. The key hurdle seems to be conceptual. We are conditioned to think of ‘organisms’ as things that are cohesive in space. But this cohesiveness is the very thing that superorganisms (should they exist) would try to avoid. If superorganisms are solving the problem of size, then their lack of (physical) cohesiveness is not a ‘flaw’. It is a design feature.
Take, as an example, a colony of ants. The colony might have the same biomass as a small mammal, but it can do things the mammal cannot. The cells of the mammal are stuck together. But the ants can spread out in search of food. As superorganisms, ants solve the tyranny of geometry. The result is that given the same concentration of food, an ant colony can grow far larger than a single mammal.
Human superorganisms
The rise of superorganisms undoubtedly marks a major transition in evolution. But has it led to a major transition of size?
The answer depends on where you look.
Ant colonies may be superorganisms, but they are not super-massive ones. (I doubt that a single ant colony surpasses the mass of a blue whale.) To find super-massive superorganisms, we need to look elsewhere. We need to look at ourselves.
An increasing number of scientists believe that human culture is a major transition in evolution. Not only are humans ultra-social — organizing in groups that deserve to be called superorganisms8 — we coordinate at scales never before seen on Earth.
Let’s have a look at this scale. Suppose we treat human groups as superorganisms. Figure 4 shows how these superorganisms fit into the biomass spectrum. Here I’ve taken US institutions (firms and governments) and treated them like organisms. Their mass is equal to the biomass of the humans they employ. And their abundance is the number of similarly-size institutions contained within the US. When we treat US institutions as superorganisms, we find something amazing: they extend the biomass spectrum by 4 orders of magnitude.

US institutions, it seems, behave like the rest of life on Earth: their abundance declines with mass. There is something eerie about this result. Humans often consider ourselves distinct from the animal kingdom. But it turns out that we are not. When it comes to size, our groups behave as if they were organisms.
The difference, however, is that our groups can become super-massive. When treated as a superorganism, the US Federal government is a creature some 10,000 times bigger than a blue whale. Were this mass packed into one animal, the beast would be about three quarters the height of the Empire State Building.9 Unfortunately, such a behemoth would be quickly reduced to a puddle of guts.
Although there will never be a real-life Godzilla, there may already be a Godzilla-sized superorganism. It is us.
Revolutions of size
In the 4 billion years since it emerged, life has evolved to mammoth proportions. Along the way, it repeatedly ran into barriers caused by the tyranny of geometry. But each time, life found ways of sidestepping these limits.
The key solution seems to be sociality.
In 3 separate revolutions, life used sociality to get bigger. Figure 5 gives a birds-eye view of these transitions. First, prokaryotes went social, creating eukaryotes. Then eukaryotes went social, creating multicellular organisms. Finally, multicellular organisms went social, creating superorganisms.
As we humans peer down from the top of life’s size pyramid, it is tempting to gloat. But we should remember that life at the top is precarious. When a mass extinction strikes, big organisms are the first to go.
On that front, we tend to think of evolution as a steady march towards greater size and complexity. But there is nothing inevitable about this trend. In the distant future, it will likely reverse. As the sun ages, it will heat up, bathing the Earth in more radiation. The result will be a gradual march towards simplicity and asociality. Multicellular organisms will slowly be killed off, followed by single-celled eukaryotes. Eventually, the planet will look much like how it started: a world of autonomous prokaryotes eking out a microscopic living.10
Of course, this future is so distant that it’s not worth worrying about. We have more imminent problems on our hands. Human superorganisms may be the pinnacle of evolution (to date). But by feeding these superorganisms fossil fuels, we may have inadvertently sown the seeds of our own destruction. Humans live at a unique point in Earth’s history — a period when life has had time to grow complex, but not yet been killed off by the heating sun. Perhaps we should not squander the opportunity.
taken from here